
Basidiomycetes. Royas y carbones
Por: Juan Luis Menéndez
Características generales
El grupo de los teliomycetes está formado por hongos parásitos de plantas comúnmente denominados royas y carbones, caracterizados por la producción de esporas de resistencia, con una pared gruesa, llamadas teliosporas. Es en estas esporas donde ocurre la cariogamia y su germinación origina un promicelio o tubo germinativo al cual migra ese nucleo diploide. Tras la meioisis se forman las basidiosporas, haploides, ya sea directamente sobre el promicelio o en el ápice de pequeñas evaginaciones que surgen de él. Estas basidiosporas germinan originando o bien hifas que producen otros tipos de esporas indirectas o bien conidios. No forman por tanto cuerpos fructíferos o basidiomas. Los poros septales son casi siempre simples, siendo muy raro la presencia de doliporos con parentosomas perforados, típicos del resto de basidiomycetes.Uredinales. Las royas

Puccinia ligustici";?>
Cuando la espora de una roya alcanza la superficie de una planta susceptible de ser parasitada, ocurren una serie de fenómenos que conllevan la penetración e instalación del micelio del hongo dentro de la planta. Una vez en el interior, el micelio crece entre las células y para obtener su alimento desarrolla unoas estructuras llamadas haustorios. Cuando una hifa intercelular se encuentra con una célula su porción terminal se delimita mediante un septo; de esta célula terminal, o célula madre del haustorio, se forma un botón de perforacion que perfora la célula de la planta arrastrando su membrana plasmática, a la vez que el botón comienza a hincharse; en ningún momento se rompe la membrana de la célula de la planta, si no que haustorio y célula permanecen separados por una pequeña región delgada conocida como vaina.
El ciclo de vida de los Uredinales
El ciclo de vida de las royas es una serie de sucesos muy variable y compleja. Todas las royas, excepto las denonimadas imperfectas, producen teliosporas, ya que se considera a la teliospora la fase perfecta al ser en la que se producen los fenómenos de cariogamia y meiosis; además de las teliosporas, las royas producen además otros tipos de esporas que van alternando en el ciclo, que puede tener hasta cuatro o cinco fases. Así, las fases del ciclo han quedado establecidas en:- Fase 0: espermogonios portadoras de espermacios e hifas receptoras.
- Fase I: ecio o ecidio portadores de eciosporas o ecidiosporas.
- Fase II: uredinio portadores de urediniosporas.
- Fase III: telio portadores de teliosporas.
- Fase IV: basidios portadores de basidiosporas.
- Royas macrocíclicas: con las cinco fases reproductoras.
- Royas demicíclicas: carecen de fase uredinal.
- Royas microcíclicas: la teliospora es la única fase binucleada existente.
Espermogonios
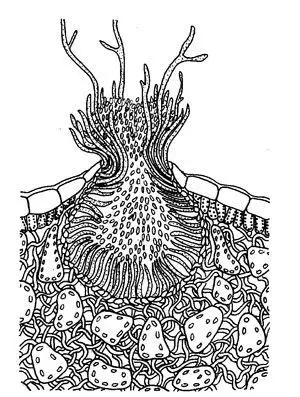
Esquema del espermogonio de Puccinia

Espermogonios y espermacios de Gymnosporangium en hojas de Cotoneaster.
En un recuadro se puede ver como segregan una gota de néctar. Enrique Rubio
Ecios y eciosporas
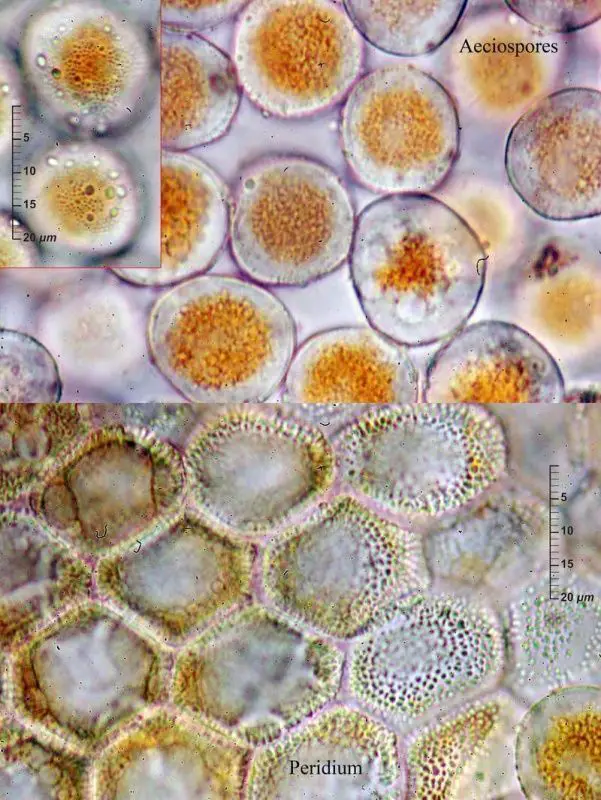
Eciosporas y pseudoperidio de Puccinia caricina,
Enrique Rubio
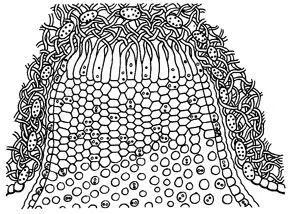
Esquema del ecio de Puccinia
Uredinios y urediniosporas
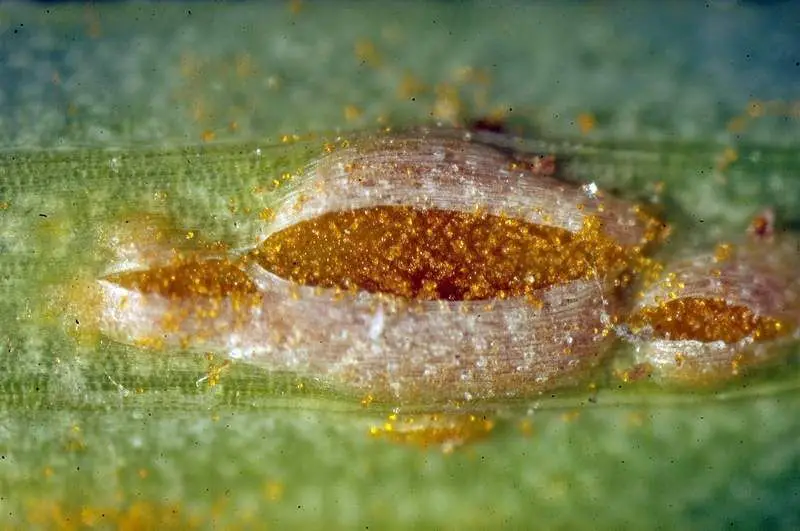
Uredosoro de Puccinia allii, Enrique Rubio
Telios y teliosporas
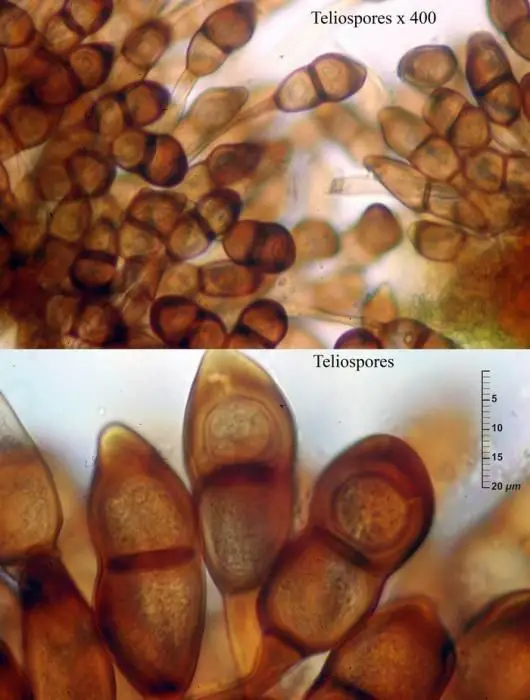
Teliosporas de Puccinia sorghi, Enrique Rubio
Basidios y basidiosporas
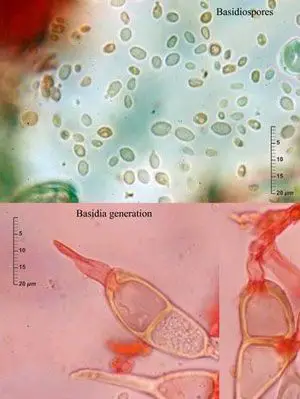
Germinación de un basidio de Puccinia annularis,
Enrique Rubio
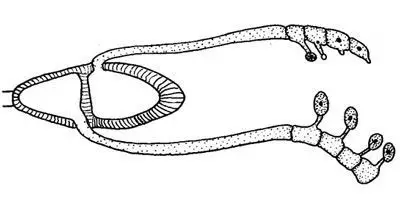
Germinación de una teliospora
Ciclo biológico de Puccinia graminis
Al finalizar el verano comienzan a madurar las teliosporas o teleutosporas sobre las hojas y tallos del trigo o cualquier otra gramínea susceptible de ser infectada; son esporas bicelulares, al inicio binucleadas. Gracias a sus gruesas paredes han permanecido aletargadas durante el invierno en estado uninuclear y diploide al haberse producido ya la cariogamia. Durante la primavera, cada célula diploide de la teliospora germina, originando un promicelio al que pasa el núcleo diploide; se produce entonces la meiosis y se forman cuatro núcleos haploides que quedan delimitados por septos, apareciendo así cuatro células, en cada una de las cuales se produce un esterigma y a cuyo extremo engrosado pasa el núcleo para formar una basidiospora. De esta forma se producen los fragmobasidios con cuatro basidiosporas cada uno, de las cuales dos son + y dos son -, de diferente polaridad.
Ecios de Puccinia graminis, Juan Luis Menéndez
Tras esta dicariotización ya se producen los ecidios, que al madurar rompen la epidermis para dejar salir las esporas; toman aspecto acopado y color anaranjado. Las eciosporas periféricas pierden su función esporal y se sueldan entre sí para formar una envoltura resistente llamada pseudoperidio; las restantes se forman en cadenas y acaban siendo dispersadas por el viento hasta que alguna cae en la hoja de una gramínea susceptible de ser infectada, como por ejemplo el trigo. Entonces germina produciendo un tubo germinativo que entra en la hoja por los estomas de la misma, desarrollando un micelio dicariótico intercelular. Al poco el micelio comienza a originar los uredinios o uredosoros, formados por mitosporas pediculadas, binucleadas y ovadas, ornamentadas con finas espinas que son las uredosoras o urediniosporas; su acumulación bajo la epidermis acaba con su rotura, quedando entonces libres las esporas, lo que exteriomente se observa como una pústula alargada de color pardo rojizo. Estas esporas extienden la infección, ya que son capaces de germinar al madurar, originando de nuevo un micelio que de nuevo producirá uredosporas. Al final, más o menos en la época de maduración de la planta, el micelio dicariótico produce las teliosporas, bicelulares, en los uredosoros u otras estructuras llamadas telios; en las células de estas esporas se produce la fusión de los dos núcleos, la cariogamia, y este estado monocariótico haploide, gracias a su pared, se prepara para el letargo invernal.
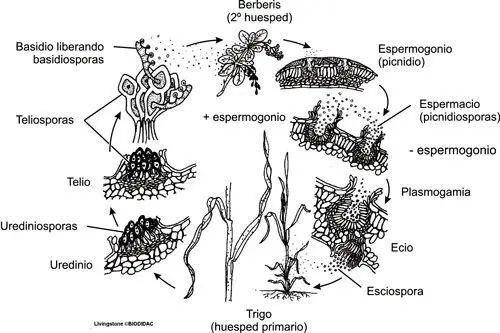
Ciclo biológico de Puccinia graminis
Heterecia en las royas
Las royas son parásitas estrictas de plantas y por lo general muy específicas, de forma que una especie de roya ataca a una misma especie de planta mientras que otras royas pueden atacar a varias especies relacionadas. Una característica muy peculiar de los uredinales es la heterecia; un hongo heteroico necesita dos huéspedes distintos para completar su ciclo, produciendo las fases 0 y I en una planta y las restantes en otra (no tienen porque estar relacionadas filogenéticamente), siendo aquel sobre el que producen su fase perfecta o telial (III) el huesped primario y el otro el huesped secundario o alternativo. Un hongo autoico es el que complementa su ciclo sobre un mismo huesped.En el caso de las royas heteroicas es necesario conocer los huespedes para concer el ciclo vital. Una de las royas más importantes, la roya del trigo (Puccinia graminis), como se ha visto es una roya heteroica cuyo huesped primario es el trigo y el secundario el agracejo producía importantes daños en los cultivos cerealistas y por más que se quitaban los cereales dañados el mal no se iba; se empezó a sospechar de la importancia del agracejo y en algunos países, como Francia, se promulgaron leyes para eliminar el agracejo de las cercanías de los campos de cultivo. De esta forma, al eliminar el huesped secundario, se rompía el ciclo de la roya y se disminuía la infección por el mismo.
Clasificación
Se han realizado muchas clasificaciones del grupo debido a la dificultad que entraña conocer el ciclo vital de las mismas. Como sucede en los ascomycetes, las fases imperfectas de las royas son más fáciles de encontrar que la perfecta (telios); realizar una clasificación basada en la morfología de telios y teliosporas implica conocer esta fase, pero en muchas ocasiones, especialmente si el ciclo es heteroico, disponer de otra fase hace prácticamente imposible conocer la especie. Por esto se ha creado el grupo artificial de los Uredinales Imperfecti, que de forma similar a los Deuteromycetes, recoge de forma temporal las fases eciales y urediniales cuyas fases perfectas se desconocen; a este grupo pertenecen los género forma Aecidium, Caeoma, Peridermium, Roestelia y Uredo:- Caeoma: formas eciales que carecen de pseudoperidio, por lo que no forman las copas características del género siguiente.
- Aecidium: formas eciales cuyos ecios tienen forma de copa, con un labio corto y erecto.
- Roestelia: el labio del ecio sobresale mucho de la epidermis y acaba por deshilacharse. Es la fase ecídica de Gymnosporangium
- Peridermium: el labio ecial es grande, conspicuo, de color pálido y emerge mucho de la superficie de la planta.
- Uredo: se agrupan aquí todas las fases uredinales.
Ustilaginales. Los carbones

Ustilago avenae
Los carbones carecen, como las royas, de cuerpos fructíferos o basidiomas y son parásitos intercelulares. Se diferencian de las royas o Uredinales en que el micelio monocariótico vive como saprófito, siendo parásito sólo el micelio dicariótico; tampoco existen en este grupo especies heteroicas, y sus ciclos vitales son más sencillos.
La plasmogamia tiene lugar por fusión de dos células compatibles cualquiera, no siendo necesario células sexuales diferenciadas como los espermacios e hifas receptoras que aparecen en las royas; así, pueden fusionarse dos basidiosporas, dos conidios o incluso dos yemas producidas por las anteriores; pueden incluso fusionarse micelios.
En los Ustilaginales, las teliosporas son las células características del grupo. Se forman en soros que si bien generalmente se desarrollan sobre las flores también pueden hacerlo en tallos, hojas, frutos e incluso raíces; en el momento de la esporulación, el micelio dicariótico se desarrolla con profusión en la planta, produce muchos septos y pasa a estar formado por hifas de células cortas. En un determinado momento el protoplasto de cada célula se redondea y sus paredes se gelifican; luego, el protoplasto segrega a su alrededor una pared gruesa que lo acaba convirtiendo en una teliospora redondeada. La ornamentación, tamaño, forma y color son caracteres necesarios para la determinación de las especies; en muchas especies son sólo células aisladas, pero en otras se agrupan formando bolas de esporas, en las que en ocasiones se pueden diferenciar células fértiles y células estériles. La germinación de la teliospora origina un promicelio, al igual que las royas, sobre el que se forman directamente las basidiosporas (también llamadas esporidios), pero sin la mediación de esterigmas, por lo que no son expulsados con fuerza. Asexualmente se reproducen por conidios producidos a partir del micelio uninucleado y del micelio binucleado; estos conidios se desarrollan sobre esterigmas y son expulsados bruscamente, de forma similar a las basidiosporas de otros basidiomycetes.
Ciclo biológico de Ustilago maydis
El carbón del maíz, Ustilago maydis, forma grandes masas pulverulentas en el maíz y es uno de los principales problemas de los cultivos de este cereal. Las teliosporas maduras, conocidas también como ustilagosporas, son unicelulares, uninucleadas y diploides. Tras pasar un posible periodo de reposo, se produce la germinación al abrirse su pared para dejar paso a un tubo de germinación (promicelio), al cual pasa el núcleo; se produce la meiosis y los núcleos generados se reparten uniformemente por el promicelio; la formación de tabiques entre ellos hace que el tubo pasa a estar formado por células uninucleadas haploides, algo que recuerda a un fragmobasidio. A continuación, cada núcleo sufre una mitosis, y uno de los dos núcleos originados pasa a una yema que se forma en cada célula, la cual por estrangulación se libera; se diferencia de una basidiospora en sentido estricto en que por cada célula del "basidio" se forman varias esporas y no una sóla, además de estar sostenida por un esterigma; estas basidiosporas se conocen como esporidios. Al ser liberados originan un micelio con aspecto de levadura, uninucleado, que vive saprófito sobre la superficie de la planta, a diferencia de los micelios de los uredinales que pasan al interior del tejido. Si sobre la misma planta se forman micelios compatibles, puede ocurrir un proceso de somatogamia entre células de un micelio + y un micelio -, origiándose un micelio dicariótico. Este micelio es el que produce la infección al penetrar y ramificarse en el interior de los tejidos, llegando por lo general a las partes reproductivas de la planta; las ramificaciones que alcanzan la superficie originan conidios binucleados, que son dispersados por el viento y así provocan nuevas infecciones. El micelio que alcanza algunos órganos (normalmente los ovarios y anteras), se convierte en intracelular, destruyendo por completo los tejidos; se forman así grupos densos de hifas que adquieren forma redondeada, su contenido se hace granuloso y sus paredes se gelatinizan; luego cada célula binucleada sufre cariogamia y se engruesan las paredes, originando así la ustilagospora inicial; estas masas densas, los teliosoros, son de color negruzco, lo cual recuerda al polvo de carbón, lo que da nombre al grupo.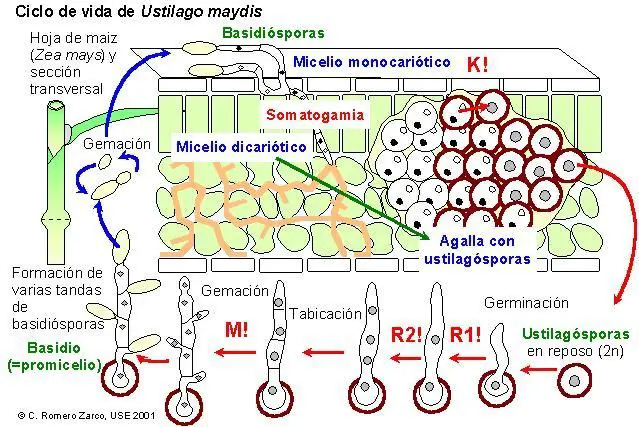
Ciclo biológico de Ustilago maydis, © C. Romero Zarco
Hashtags:
#hongos#setas#micologia#basidiomycete#ascomycete#myxomycete#protistas#liquenes#intoxicacion-por-setas
Juan Luis Menéndez
Me gusta la divulgación de la naturaleza y el patrimonio cultural, motivos por los que he decidido comenzar el desarrollo de asturnatura.com. Soy un amante de la botánica y la geología.
Artículos similares

Citar como
Menéndez Valderrey, Juan Luis. "Basidiomycetes. Royas y carbones". asturnatura.com [en línea] Num. 260, 22/03/2010 [consultado el 9/4/2024]. Disponible en https://www.asturnatura.com/temarios/biologia/hongos/basidiomycetes-teliomycetes-uredinales-ustilaginales-royas-carbones-rust.
ISSN 1887-5068