


Los ribosomas
Los ribosomas son pequeñas partículas en las cuales tiene lugar la síntesis de proteínas en los seres vivos. Son solo visibles al microscopio electrónico y fuero descritos por primera vez por Palade en 1953 como pequeñas estructuras globulares abundantes en el citoplasma de la célula.
Un ribosoma es un complejo formado por varias moléculas de RNA ribosómico asociadas a más de 50 proteínas diferentes. El rRNA constituye más de la mitad del peso del ribosoma y desempeña una función catalítica en el proceso de la síntesis proteica. Se cree que las proteínas ribosómicas incrementan la función de los rRNA. Cada ribosoma está formado por una subunidad pequeña y una subunidad grande, que se disocian al final de cada proceso de síntesis proteica. La subunidad pequeña se une al RNA mensajero (mRNA) y a las moléculas de RNA transferente (tRNA), mientras que la subunidad grande cataliza la formación de los enlaces peptídicos.Los ribosomas de las células eucariotas son de mayor tamaño que los de las células procariotas. En la siguiente tabla se muestran las diferencias entre ambos.
| Ribosoma eucariota - 80S | Ribosoma procariota - 70S |
|---|---|
| Subunidad mayor - 60S 49 proteínas RNA 5S, RNA 5.8S y RNA 28S |
Subunidad mayor - 50S 34 proteínas RNA 5S y RNA 23S |
| Subunidad menor - 40S 33 proteínas RNA 18S |
Subunidad menor - 30S 21 proteínas RNA 16S |

Diferencias entre un ribosoma eucariota y un ribosoma procariota
Localización de los ribosomas
Los ribosomas 80S de las células eucariotas pueden encontrarse libres en el citosol o unidos a la cara citosólica de las membranas del retículo endoplasmático.Los dos tipos de ribosomas del citoplasma son estructural y funcionalmente iguales, pero según estén libres o unidos al retículo endoplasmático sintetizan proteínas cuyos destinos finales son diferentes:
- En los ribosomas libres se sintetizan proteínas del citosol, núcleo, peroxisomas, mitocondrias y cloroplastos, en el caso de los vegetales.
- En los ribosomas unidos al retículo endoplasmático se sintetizan las proteínas del RE, aparato de Golgi, lisosomas, membrana plasmática y las destinadas a ser secretadas al exterior celular.

A la derecha, ribosomas libres en el citosol, agrupados en una organización casi circular llamada polisoma. A la izquierda, ribosomas unidos a la membrana del retículo endoplasmático
En las células eucariotas hay también ribosomas 70S, similares a los bacterianos, en el interior de las mitocondrias y de los cloroplastos. En estos ribosomas se sintetizan las proteínas codificadas por el DNA mitocondrial o del DNA del cloroplasto.
Origen de los ribosomas
La formación de los ribosomas comprende la síntesis y el ensamblaje de sus dos componentes, las proteínas y los rRNA. Las proteínas ribosomales se sintetizan en el citosal y los rRNA en el nucleolo. El ensamblaje tiene lugar en el nucleolo.Las células contienen múltiples copias de los genes para los rRNA para poder satisfacer la demanda de transcripción de elevado número de moléculas de rRNA que son necesarias para sintetizar los ribosomas. Por ejemplo, las células de mamífero en continuo crecimiento contienen 5 y 10 millones de ribosomas, que deben sintetizarse cada vez que la célula se divide. Las células contienen por ello múltiplas copias de los genes rRNA. El genoma humano por ejemplo contiene aproximadamente unas doscientas copias del gen que codifica para los RNAr 28 S, 18 S, 5.8 S dispuestas de manera secuencial (en tándem) con un DNA espaciador que no se transcribe separando cada unidad repetida en cinco cromosomas humanos diferentes (13,14,15,21,22) y aproximadamente 200 copias del gen que codifica para el RNAr 5S en el cromosoma 1.
Síntesis y procesamiento de los RNA ribosómicos (rRNA)
Los RNAr nucleolares 18 S, 5.8 y 28 S son sintetizados (transcriptos) por la RNA polimerasa I a partir de los genes (DNAr) que codifican los RNAr. Lo que permite que la transcripción se pueda visualizar fácilmente con microscopia electrónica, cada uno de los genes de ARNr están colocados en serie y se encuentran rodeado de ARN en crecimiento densamente empaquetados, (unidos a diferentes proteínas de procesamiento y ribosómicas) dando lugar a estructuras en forma típica de "arbol de navidad".El transcripto primario de los genes RNAr es un pre-RNAr (47S en células de mamífero) de gran tamaño que contiene los RNAr 5.8 S, 18S y 28S, dos espaciadores externos (ETS) que también son transcritos localizados en los extremos 5´ y 3´del pre-RNA y dos espaciadores internos (ITS) que se sitúan entre las secuencias de los RNAr 18s, 5.8 s y 28s. Así la estructura del pre-RNAR es: 5´-ETS-18S-ITS-5.8-ITS-28S-3´.
Mediante escisiones sucesivas (realizadas por endonucleasas específicas) de este transcrito primario se produce la liberación de los RNAr 5.8 S, 18S y 28S. Este procesamiento del pre-RNAr requiere de la intervención de un numerosas grupos de proteínas (unas 300) y RNAs localizados en el nucléolo, denominados RNAs nucleolares pequeños (RNAsno), los cuales al unirse a proteínas constituyen unas partículas denominadas proteínas ribonucleares pequeñas (abreviadamente RNPsno). Cada RNPsno está constituida por un único RNAsno asociado a ocho o diez proteínas. Las células humanas contienen aproximadamente 100 especies diferentes de snoRNP Por ejemplo la RNPsno llamada U3 es necesario para la escisión inicial del pre-RNAr que se produce en la ETS 5. De manera similar el RNPsno U8 provoca la escisión del pre-RNAr en RNAr 5.8 S, 18S y 28S, mientras que la RNPsno U22 es responsable de la fragmentación adicional de pre-RNAr para dar lugar al RNA 18 S.
Para alcanzar la madurez funcional los RNAr sufren una extensiva modificaciones covalentes, metilaciones de ciertas bases nitrogenadas y de residuos de ribosa (los grupos hidroxilos 2' (2'-O-metilación) o la conversión de uridina en pseudouridina. Estas modificaciones que requieren de la actividad de las RPNsno, la mayoría de los RNAsno contienen secuencias cortas de 15 nucleótidos que son complementarias a las secuencias de los RNAr 18s y 28s que sirven para reconocer, seleccionar (e.g.los sitios de metilación) y dirigir a las enzimas que catalizan las modificaciones al sitio adecuado de las secuencias del pre-RNA. En las células animales el procesamiento de pre-RNAr 47S implica la metilación de aproximadamente cien restos de ribosa y 10 bases, además de la formación de cien pseudouridinas. La mayoría de estas modificaciones ocurre durante o inmediatamente después de la síntesis de pre-RNAr aunque algunas tienen lugar en etapas posteriores del procesamiento del pre-RNAr.

Esquema de la síntesis y ensamblaje de los ribosomas
Ensamblaje de los ribosomas
Los RNAr maduros 5.8 S, 18S y 28S y el RNAr 5S se combinan en el nucleolo con las proteínas ribosómicas (importadas desde el citoplasma) para formar las subunidades ribosomales pre- 40S y pre-60S. Estas pre-subunidades son exportadas a través de los complejos del poro nucleares (NPCs) al citoplasma donde se termina la maduración.Los genes que codifican para las diferentes proteínas ribosomales se transcriben fuera del nucleolo por la RNA polimerasa II, originando RNAm que son transportados a través de los NPCs al citoplasma donde son traducidos en proteínas ribosomales en los ribosomas citoplasmáticos. Las proteínas ribosomales son transportadas entonces de nuevo a través de los NPCs al nucleolo donde se ensamblan con los RNAr maduros para formar las partículas pre-ribosómicas.
La asociación de las proteínas ribosomicas con los RNAr tiene lugar a lo largo de la síntesis y procesamiento del pre-RNA. La maduración de la pre-subunidad mayor 60S sigue una ruta diferente de la menor 40S. La maduración de la subunidad pequeña que solo contiene RNAr 18S es más sencilla e implica cuatro escisiones en le pre-RNA 47S. La escisión final de la que resulta el RNAr 18S se produce tras el transporte de la subunidad 40S al citosol, mientras que la maduración de la subunidad 60S que contiene implica multiples escisiones del pre-RNA en el núcleo y se completa totalmente dentro del nucleolo. Por lo tanto la mayoría de las particulas preribosómicas del nucleolo son precursores de las subunidades grandes 60S. Las etapas finales de la maduración de los ribosomas siguen a la salida de las partículas preribosomales al citoplasma, formando las subunidades ribosómicas 40S y 60S maduras funcionalmente capaces de formar los ribosomas 80S encargados de llevar a cabo la síntesis de proteínas celulares.
Función de los ribosomas
Los ribosomas intervienen en la síntesis de proteínas, en un proceso llamado traducción y que se estudiará con más detalle en el capítulo correspondiente.El ribosoma se une primero a un punto específico de la molécula de RNA mensajero (mRNA), sobre la que se va desplazando y va traduciendo la secuencia de nucleótidos a la secuencia de aminoácidos de la proteína. Cuando un ribosoma llega al final del mensaje, la proteína reción sintetizada se libera y las dos subunidades del ribosoma se disocian del mRNA quedando libres en el citosol.
Generalmente varios ribosomas traducen simultaneamente la misma molécula de mRNA dando lugar a un polisoma o polirribosoma.

Esquema de la traducción, en la que en el ribosoma se traduce la información contenida en el RNA mensajero en una secuencia de aminoácidos que formarán una proteína

Comparte en:
Índice:
Más populares en Bioquímica:
 Fases de la Meiosis
Fases de la Meiosis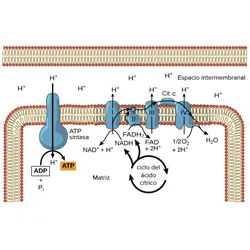 La cadena de transporte de electrones y fosforilación oxidativa
La cadena de transporte de electrones y fosforilación oxidativaPalabras clave:
Artículos similares



Citar como
Aida Lorenzo Corchón. "Los ribosomas". asturnatura.com [en línea] Num. 502, 10/11/2014 [consultado el 24/4/2025]. Disponible en https://www.asturnatura.com.
ISSN 1887-5068