
Morfología de los trilobites
Por: Juan Luis Menéndez
Esquema general del cuerpo
Vimos en el capítulo anterior las principales partes corporales de los trilobites, el céfalon, el tórax y el pigidio. En este capítulo veremos con detalle cada una de estas partes. En las dos figuras siguientes se muestra con más detalle las principales características de la morfología dorsal (figura 1) y ventral (figura 2) de un trilobite.
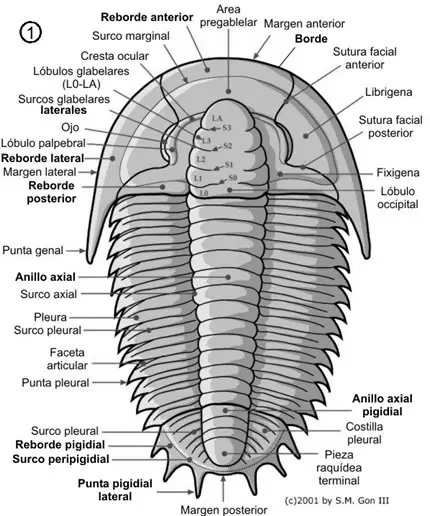
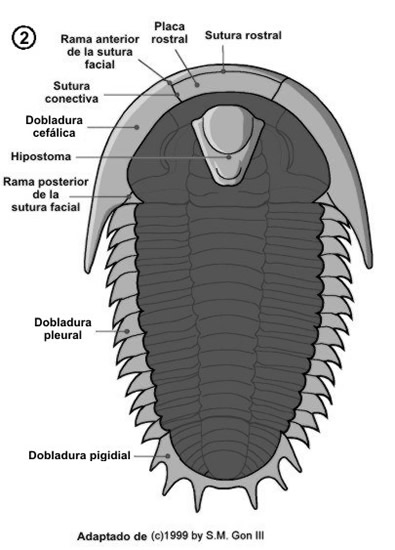
Este esqueleto se encuentra atravesado por numerosas sedas con función sensitiva y presenta numerosas ornamentaciones crateriformes o tuberculosas.
Región cefálica o céfalon
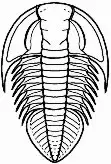
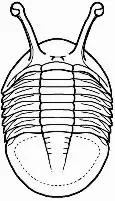
En el céfalon la parte que más destaca es la glabela, que no es más que la parte anterior del lóbulo axial, convexa y que presenta un número variable de surcos transglabelares o surcos laterales que corresponden al número de segmentos que forman esta región. En todos los trilobites el céfalon es una placa formada por un número variable de segmentos fusionados en los que se encuentran los ojos, las suturas faciales y el hipostoma, y la gran variedad de morfologías que puede adoptar son en la gran mayoría de las especies caracteres identificativos.
En el esqueleto externo del céfalon se pueden diferenciar un cranidio, formado por la glabela y las mejillas fijas o fixígenas; lateralmente al cranidio están las librígenas o mejillas libres, separadas del mismo por la sutura facial de la que posteriormente trataremos (ver fig. 3). Las mejillas corresponden a las regiones pleurales cefálicas.
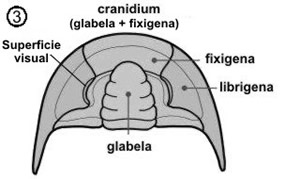
En el céfalon se dividen zonas atendiendo a las suturas, placas y posición que ocupan de gran importancia en la determinación de las diferentes especies (ver fig. 4).
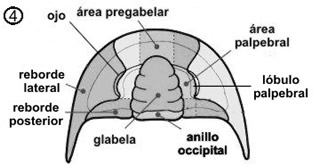
La glabela varía considerablemente entre las especies, tanto en forma como en estructura, siendo en unos casos bastante pronunciada con lóbulos laterales o en otros menos conspicua. Se cree que el estómago se encontraba entre ella y el hipostoma de la parte inferior, por lo que su tamaño guardaría una estrecha relación con el mismo y por consiguiente con la dieta de la especie. Variaciones en su tamaño y el número de estrías pueden considerarse indicadores del ambiente y dieta.
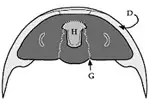
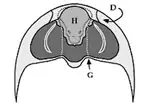
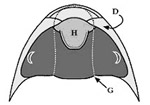
El hipostoma (fig. 2), situado en la zona ventral de la región cefálica, es una pieza de exoesqueleto que se cree que formaba parte de la boca. Su borde anterior se encuentra bajo el borde anterior de la glabela por lo general. Puede adoptar gran variación morfológica usada como caracter identificatorio y básicamente se diferencian 3 tipos:
- Flotante: se encuentra separado y libre del pliegue anterior y su borde anterior coincide con el de la glabela.
- Conectado: se encuentra fijo al pliegue anterior de la placa rostral y su borde coincide con el anterior de la glabela.
- Desplazado: semejante al anterior, pero el borde anterior no coincide con el de la glablela.
Flotante
Conectado
Desplazado
El hipostoma flotante muestra diseños bastante conservadores a lo largo de su evolución; su forma suele ser ovoide y carente de extensiones u ornamentaciones. La alimentación generalista a base de partículas es la causa de las formas tan simple y poco variadas de su hipostoma.
Por el contrario, los otros dos modelos han sufrido una mayor diversificación que originó una serie de modelos muy variados en forma y tamaño que ha servido para clasificar muchas especies de trilobites y especular sobre sus hábitos alimentarios. Se encuentran en especies que se cree que fueron depredadoras, por lo que su anclaje a la placa rostral proporcionaría más solidez al mismo y ante los posibles ataques a las presas; la especialización de formas estaría relacionada con diferentes tipos de presas o comportamientos alimenticios.
Las suturas están relacionadas con las mudas. No hemos de olvidar que los trilobites son artrópodos y como tales tienen un esqueleto externo; el crecimiento del animal viene entonces marcado por las mudas de este exoesqueleto, que debe romper por unas líneas fijas (las suturas) para salir con un nuevo exoesqueleto de mayor tamaño. Estas suturas indican el lugar de la región cefálica por donde se separan las placas para permitir la salida del animal mudado y por lo general discurren desde el borde anterior del céfalon, alcanzan la superficie visual, separando la superficie visual el lóbulo palpebral, y continúan hacia el borde posterior de la misma. Hay tres tipos principales de suturas faciales en función del lugar donde finalizan en la zona posterior (fig. 5):
- Propárica: el final se encuentra en el borde anterior al ángulo o espina genal.
- Gonatopárica: el final de la sutura desemboca posteriormente al ángulo genal o muy próximo a él.

La sutura facial además parte la zona dorsal de la mejilla en las dos que vimos anteriormente, la fixigena y la librigena; la segunda, la mejilla libre, se desarticula durante la muda y también frecuentemente tras la muerte del animal, por lo que es fácil encontrar fósiles sin esta pieza.
Las suturas dorsales no finalizan en los margenes laterales, sino que se extienden por la región ventral de la zona cefálica a lo largo de la dobladura cefálica, la placa rostral y el hipostoma.
En el céfalon también se encuentran los ojos. Los trilobites fueron los primeros seres vivos en desarrollar un sistema visual y éste es bastante semejante al que tienen los crustáceos o insectos actuales, los ojos compuestos, formados por múltiples facetas independientes cuyos campos visuales individuales procesados por el cerebro forman la imagen completa. Fueron además los primeros en desarrollar este tipo de ojo. Se encuentran el borde externo de la fixigena, a ambos lados de la glabela y adyacentes a la sutura facial. Hay grupos de trilobites que carecen de ojos (Agnostina), mientras que otros los pierden tras iniciar una reducción de su tamaño de forma secundaria y otros sufren el proceso contrario, desarrollando ojos de gran tamaño en función del ambiente en el que evolucionaron.
En los vertebrados es posible que la lente ocular o cristalino varíe su forma para lograr enfocar; sin embargo, en los trilobites y demás artrópodos las lentes son rígidas, por lo que no pueden modificarse para enfocar. Para conseguirlo disponen de una segunda lente que combinada con la anteiror soluciona el problema del foco.
Hay tres tipos de ojos en los trilobites:
- Holocroal: presente en la mayor parte de los órdenes de trilobites, están formados por hasta 15000 facetas pequeñas. La membrana de la córnea cubre sólo la superficie de las facetas que están en contacto las unas con las otras.
- Esquizocroal: este tipo se considera una innovación presente en el grupo de los Phacopida. No tiene tantas facetas como el grupo anterior, solamente alrededor de unas 700, pero su tamaño es mayor. Cada una de las facetas tiene una córnea individual y se encuentran separadas por una capa esclerotizada relativamente gruesa; la membraba de la córnea penetra en la parte interna de la faceta y se llama en esta zona membrana intrascleral.
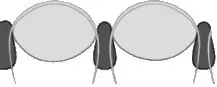
- Abatocroal: solamente se han encontrado en el género Eodiscina del Cámbrico. Sólo tiene unas 70 lentes o facetas de pequeño tamaño, cada una con una córnea individual y separada del resto de facetas por una capa esclerotizada no tan gruesa como la presente en el modelo ocular anterior. La córnea no penetra en el interior de la faceta.

Estos ojos se encuentran en número par y en posición dorsal, generalmente vertical para lograr una visión completa del entorno, en muchos casos de 360º. Muchos trilobites presentan además dos ojos simples u ocelos conocidos como máculas, en posición ventral y en el hipostoma, que podrían semejar a los ocelos de las larvas nauplius de los artrópodos y que no desaparecerían durante la ontogenia, al igual que ocurre en otros muchos artrópodos actuales.
Entre los ojos de los trilobites existe una gran variedad morfológica que responde a las diferentes adaptaciones que mostraban a los diferentes nichos en los que se encontraban. Muchas de las especies más primitivas tenían ojos con forma de media luna; el aumento de estos hasta adoptar una forma casi cónica en sección (ojos esquizocroales) permitió aumentar el campo visual, algo que en géneros nadadores como Opipeuterella hizo que sus enormes ojos le proporcionaran una visión de 360º. Especies bentónicas como las del género Cryptolithus tenían los ojos bastante reducidos o carecían de ellos, y otras que vivían en zonas con detritos o vegetación podían mostrar los ojos levantados del cuerpo sobre pedúnculos para lograr una mejor visión de los alrededores como ocurre en Neoasaphus. Especies de aguas profundas, donde no había luz, los ojos no les proporcionaban ninguna ventaja por lo que fueron reduciéndose secundariamente hasta poder acabar por desaparecer.
Polypleuraspis
Opipeuterella
 Neoasaphus
Neoasaphus
Polypleuraspis
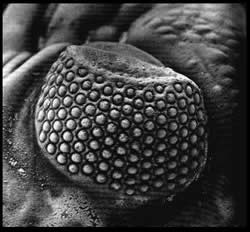
Ojo esquizocélico de Phacops
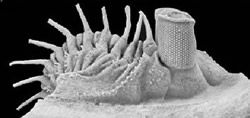
Erbenochile erbeni, encontrado en el Devónico de Marruecos en 2003, que muestra un extraordinario desarrollo ocular.
En el céfalon también se encuentran un par de antenas que surgen de los laterales del hipostoma y que en la mayoría de los casos se pierden durante la fosilización debido a su extrema fragilidad.
Tórax
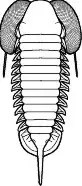
El tórax, que representa por lo general la mayor parte del cuerpo, está formado por un número de segmentos continuados unos con otros de forma flexible que permite el enrollamiento del mismo como mecanismo defensivo. Generalmente hay entre 8 y 15 segmentos, pero pueden variar entre 2 y 44. Todos ellos están formados por placas imbricadas y artículadas en la parte anterior mediante una estructura conocida como semianillo articulante y que permitía el enrollamiento del trilobite como mecanismo defensivo.
La parte ventral del tórax se caracteriza por la presencia de los apéndices, que como característica primitiva presentan las dos ramas típicas de los apéndices birrámeos, un endopodio interno y un exopodio externo (fig. 6).
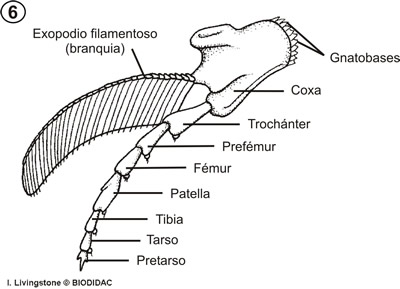
El endopodio está formado por varios artejos y el exopodio por una lámina filamentosa, que en algunos casos se emplea en la natación. El endopodio tiene función locomotora. Todos los apéndices presentan en la parte interna de la coxa, el artejo basal, y en ocasiones en todos los segmentos del endopodio, una serie de dientes más o menos agudos cuya función es triturar la comida antes de ser llevada a la boca.
En muchos grupos el tórax puede estar dividido morfológicamente en un protórax anterior y un opistotórax posterior.
Pigidio
El pigidio es la parte final del cuerpo y está formado por numerosos segmentos fusionados en una única placa. Puede variar mucho de tamaño y forma, por lo que se usa para la clasificación esta variación. Así se pueden diferenciar cuatro tipos de pigidio en relación con la región cefálica:
- Micropigido: el pigidio es mucho más pequeño que la región cefálica.
- Subisopigio: es algo menor que la región cefálica.
- Isopigido: tanto el pigidio como la región cefálica son del mismo tamaño.
- Macropigido: el pigidio es mayor que la región cefálica.
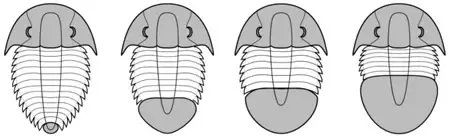
micropigido -
subisopigido -
isopigido -
macropigido
Todos los esquemas © S. M. Gon III

Juan Luis Menéndez
Me gusta la divulgación de la naturaleza y el patrimonio cultural, motivos por los que he decidido comenzar el desarrollo de asturnatura.com. Soy un amante de la botánica y la geología.
Artículos similares


Citar como
Menéndez Valderrey, Juan Luis. "Morfología de los trilobites". asturnatura.com [en línea] Num. 105, 11/12/2006 [consultado el 4/4/2024]. Disponible en https://www.asturnatura.com/temarios/geologia/trilobites/morfologia.
ISSN 1887-5068